إن ما يجمع:
المعجنات، كثير من الحلويات، أنواع الخبز المختلفة ...الخ، مادة سحريّة هي طحين
القمح.
من هنا، تأتي ضرورة دراسة تطور هذه النبتة المتواضعه المسماة: قمح.
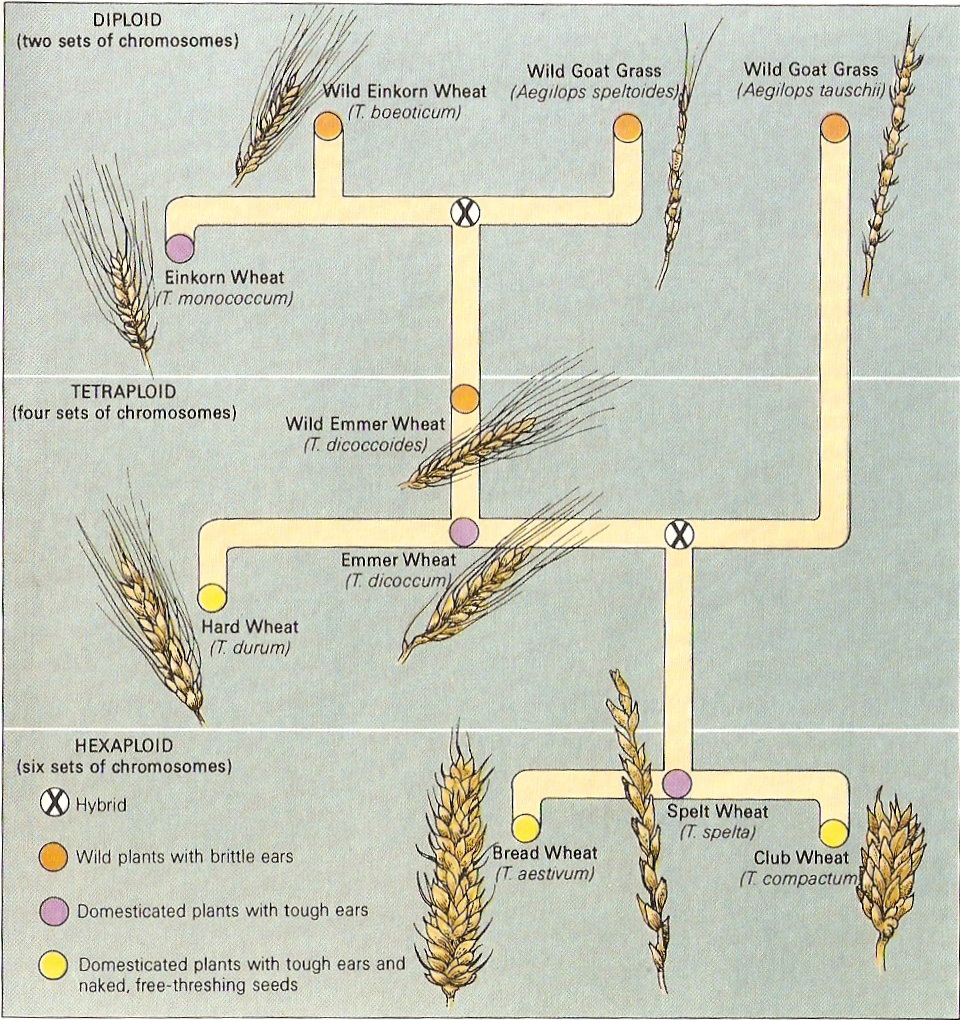
سلف متواضع وشبكيّ
ينتمي القمح الزراعي بشكل خاص إلى نوع القمح الطري Triticum aestivum
ذو
الأصل الحديث نسبياً، حيث توجد منه سلالات عديدة ويُزرَع على نطاق واسع بكل أرجاء العالم. القمح، مثل الذرة، الأرز، الشعير ...الخ، فهو نبتة تنتمي إلى الفصيلة النجيليّة، ذات دورة حياة سنويّة وتُنتِجُ البذور.
لكن، لماذا هي ذات
أهميّة كبيرة من وجهة نظر تطوريّة؟

سنابل مليئة
من نوع القمح المزروع الطري
حسناً، يتوجب البحث عن لغز هذه النبتة
في جينومها.
لا تُصاب الذعر!!
تعرف المؤسسة العلمية جيداً ما تقول،
بكلمات أخرى، هي عبارة عن نبتة تمتلك جينوماً متشكلاً من جينومات عديدة مختلفة.
نعم،
كما تقرأ بالضبط!!
في هذه الحالة، جينوم القمح الطري ذو حجم بقيمة 16000 مليون زوج من القواعد. هو رقم ضخم، سيما إذا أخذنا جينومنا كبشر بعين الإعتبار وفيه 3000 مليون زوج قواعد فقط، مُرتَّبة وفق 22 زوج من الكروموزومات
الجسديّة وزوج إضافي يحدّد الجنس.
من جانبه، يتكوّن جينوم هذا القمح من 21 زوج من الكروموزومات التي يمكن ان تُصنّث وفق الترتيب: .AABBDD

النمط النووي لجينوم قمحنا الطري
اعتباراً من هنا، يأتي ما هو ممتع
فعليّاً.
لدينا في الاعلى رسم (فيه كل الكروموزومات المرتبة وفق الحجم والتشكيل) لنوع القمح الطري، كما يمكننا ملاحظة كل الجينوم المرتّب في ثلاث
كُتل، هي: A ، B وD.
توافق الكتلة D مجموعة
الكروموزومات DD المعروفة جيداً. تشكّل، تلك الكتلة
من الكروموزومات، قسماً من جينوم القمح، وهي ذاتها عبارة عن جينوم كامل لنوع نباتيّ آخر
هو دوسر الطوشي، وهو عبارة عن عشبة تنمو بشكل برّي بصيغة دغليّة (من أدغال)
من أصل شرق متوسطي.

سنابل نوع دوسر الطوشي
من جانبهما، تشكل كتلتا كروموزومات A وB مجموعة
الكروموزومات AABB
التي بدورها، تشكّل، عملياً، جينوم نوع نباتيّ آخر، هو القمح الصلب أو قمح المعكرونة Triticum turgidum وينتمي للفصيلة النجيليّة ومن أصل شرق متوسطيّ، حيث تتم زراعة
بعض أنواعها، للآن، في تلك المنطقة من العالم.
هكذا وبناء على تلك المعطيات، نوع القمح الطري، في الواقع، عبارة عن نوع متشكِّل
من اندماج جيني وراثي لنوعين نباتيين، هما دوسر الطوشي ( كنوع دغليّ ونوع "من القمح البرّي") والقمح الصلب، اللذان باتحادهما يؤصِّلان (يُعطيان الأصل) لنوع جديد بشكل كامل من
القمح.

سنبلة القمح الصلب
مع ذلك، للآن لم تنتهِ القضيّة، فكما
رأيتم، يتحدّد جينوم نوع القمح الصلب بكتلتين AABB، لكن لماذا؟
حسناً، لقد أثبت العلماء عبر أبحاثهم بأننا بمواجهة جينومين أكثر.
فمن
جانب، لدينا مجموعة كروموزومات AA التي تنتمي
للعشبة البريّة المسماة علمياً القمح الأحمر البرّي Triticum urartu، من أقارب
"القمح"، "النشويات" وغيرها.
من جانب آخر، مجموعة كروموزومات BB هي موضع جدل. حيث توجد صعوبات بتحديد النوع الملموس الذي ينتمي له جينوم BB، توصّل بعض
الباحثين لتصنيفه بنطاق مجموعة نباتية تسمى "جماعة الدوسر Aegilops"، والتي
انتشرت على الصعيد العالمي عبر تشكيلة متنوعة جداً. ولم يطل المقام، حتى تظهر
دراسات جزيئية حديثة، تُشير لتحدُّر مجموعة الكروموزومات BB من نوع محدد من
تلك الجماعة النباتيّة هو دوسر شبه مكنسي Aegilops speltoides، عبارة عن نبتة
متوطنة في جنوب شرق أوروبا وغرب آسيا. لكن، لا تنتهي
القضية هنا، فاحتمال كبير أن الدوسر شبه المكنسي، عبارة عن نوع قد أعطى
الأصل عبر صيغة تعدد الصيغ الصبغية لنوع آخر من
"القمح البري" هو القمح البري الأرمني Triticum araraticum.
ثمّة تناقض ظاهريّ، فالرؤية الشعبية
للجنس الدوسر أبعد ما تكون عن القمح.
يُطلق بالانكليزية عليه اسم goatgrass، والذي يعني
حرفياً "عشبة الماعز" . حيث انها تمتلك فائدة وحيدة، تتمثّل
باستخدامها كمرعى للقطيع. تمّ التعرّف عليها بوصفها واحدة من آباء قمحنا.

الى اليسار سنابل نوع النشويّ البرّي نوع القمح الأحمر البري
الى اليمين سنابل نوع الدوسر شبه المكنسي البري

أصل وتطور الأنواع المختلفة من القمح
بالتالي، بذات الصيغة السالفة الذكر،
فنوع القمح الصلب، بدوره، هو نوع آخر يأتي جينومه من
اتحاد جينومين لنوعين مستقلين، يمكن ان يُعثر عليهما كأعشاب برية ومزروعة في موطنها شرق المتوسط .

التطور والتآلف اللذان أعطيا الأصل
لقمحنا
ربما لا يمكن تفادي
السؤال، كيف يمكن ان يحدث هذا؟
كمثال، نعرف بأننا لو نقم بتهجين حمار وفرس سنحصل
على بغل، وهو حيوان مصاب بالعقم كليّاً.
يتأسّس الحلّ على واقع أنّ في مجموعات
نباتية كثيرة، لا تعمل هذه الصيغة (وهذا، أيضاً، سبب صعوبة تصنيفات نباتية لجماعات
نباتية ملموسة في كثير من الأحيان). ففي هذه الحالة، يحدث الآتي:
اولاً: تشكل النجيليات نباتات تمتلك
تكاثر هجينيّ بواسطة الرياح، حيث يبدو غريباً وصول غبار طلع لنوع إلى زهور أنثوية لنوع آخر.

ازهار نوع القمح الطري، لاحظوا تموضع الأسدية (جمع سداة، العضو الذكري
في الزهرة) التي تسهل نثر غبار الطلع بواسطة الرياح
ثانياً: لسبب ما، تكون النباتات
أكثر سلاسة وراثياً جينياً من الحيوانات، بصيغة تُمكِّنها من اعطاء ظواهر متعددة
المجموعات الصبغية وحتى لتعدد مجموعات صبغية بين انواع مختلفة. حيث نرى بأن 30 نوع
مُكتشف بين جنسي البري الأحمر والدوسر: تُعرَفُ نسبة 75% منها على الأقل ناتجة عن اتحاد نوعين أو ناتجة عن اتحاد 3 أنواع.
ثالثاً: النباتات ليست هجائن
بسيطة دوماً، حيث يجب أن يحضر لدى الهجين الصِرْف نصف جينوم من نوع ونصف آخر من نوع آخر، ومن المعتاد حدوثه لدى الحيوانات، إما تُصاب بالعقم أو ذات خصوبة متعرضة للأذى.
تختلف هذه الحالة، هي ظواهر تعدّد للمجموعات الصبغية (تتضاعف المادة
الوراثية لدى أحد تلك الهجائن)، وبهذا، نمتلك جينومات كاملة وظيفيّة لأنواع محددة. فلدى الأفراد الناتجين، بالعموم، أكبر قدر من الخصوبة والقوة الفيزيائية.
في الواقع، إنتشار ظاهرة تعدد
المجموعات الصبغية الغيريّة عامة أكبر مما هو مُنتظر أو مُتوقَّع، وهي قادرة على
المساهمة في نشوء سريع للأنواع بين النباتات، ويتسبب هذا بحدوث تغيرات هامة في تنظيم
ونشاط المادة الوراثية. بكلمات أخرى، هي قوّة فعّالة محرّكة للتطور.
قواعد تعدد المجموعات الصبغية
ليست صعبة على الفهم. ففي الجدول التالي، تتوضَّح بشكل جيد:

جدول يلخص تعدد أو تغاير المجموعات
الصبغية
لنفترض وجود نوعين، نوع A ونوع B ، كلاهما متعدد الصبغيات، ما يعني، يمتلك كل واحد منهما مجموعه مضاعفة
من كروموزومات (2n) داخل خلاياه، وهو في النوع A: AA، وفي النوع B: BB . خلال التكاثر
الجنسي، يُنتج خلايا متخصصة للتناسب مع مجموعة وحيدة من الكروموزومات (n) لكل واحدة منها،
تستقبل تلك الخلايا اسم الأمشاج (عند البشر هم البويضات والحيوانات المنوية)،
بهذا، لدى النوع A أمشاج A والنوع B أمشاج B.
ففي الإلقاح
العادي بين أعضاء من ذات النوع، يتحد مشيج ذكري (n) مع مشيج أنثوي (n) مُعطياً لاقحة (2n). لكن، خلال عملية التهجين، ما يحصل هو اتحاد مشيج نوع (A) مع مشيج نوع
آخر (B) ما يؤدي لظهور
لاقحة هجينة فيها مجموعة وحيدة من الكروموزومات لكل نوع (AB).
تلك الهجائن عند الحيوانات، من النادر
امتلاكها لمستقبل.
لكن، عند النباتات، فمن الشائع بلوغها عمر التكاثر، رغم إصابتها بالعُقم في أغلبية المرات، أو مصابة به تقريباً. مع هذا، يمكننا
الآن الإلتفات الى ظاهرة معتادة في الطبيعة هي الطفرات.
لكن، بحالتنا، لا يتم الحديث عن أيّة
طفرة بعينها، بل الحديث عن الطفرة كآلة جينية وراثية، هي الأكثر بهيميّة التي يمكن
تخيلها. تضاعف كليّ وشامل للمادة الوراثية. يمكن حدوثها عبر التلوث الكيميائي
بمركبات مثل الكولشيسين؛ عبر أخطاء خلال
نشوء إنفصال الكروموزومات أثناء الإنقسام الإختزاليّ أو المُنصف، أو بسبب ثغرات
في الإنقسام الخليوي. والنتيجة أمشاج بترتيب جيني 2n أو في حالة
الهجائن مع ترتيب جيني AB.
كيف يؤصّل (يُعطي الأصل) هذا لمتعدد
أو متغاير المجموعه الصبغية؟
حسناً، تقتضي الخطوة التالية ظهور واحدة من
"اللاترجيحات الكونيّة"، تستلزم حادث تخصيب بين أمشاج ثنائية المجموعة
الصبغية.
هكذا، مشيج ثنائي (2n) + مشيج ثنائي آخر (2n) سيعطي المجال
لظهور = لاقحة متعددة المجموعات الصبغية (4n).
في حالة الهجائن، النتيجة عبارة
عن كائن: AB + AB =
AABB، وهؤلاء الأفراد بعكس الهجين
الذي أصَّلهم (أصّل، يؤصِّلُ .. يُعطي الأصل لكائن حيّ أو لشيء ..الخ) هي أنواع فعّالة وذات خصوبة جيّدة.
بكلمات أخرى، هي أنواع كاملة وجديدة وحرّة ومستقلة.
لهذا، فقمحنا هو أحد الأمثلة الأوضح عن التطور، الذي يمكننا العثور عليه، كذلك، من خلال التغيرات المورفولوجية (التشكُّلية) والوراثية
الهامة. من جانب آخر، على الرغم من كون نموذج نشوء أنواع القمح غير مألوف كليّاً في
المملكة الحيوانية؛ فإنّ تعدد المجموعات الصبغية هو ظاهرة عادية في المملكة
النباتيّة.
علم الوراثة والتدجين
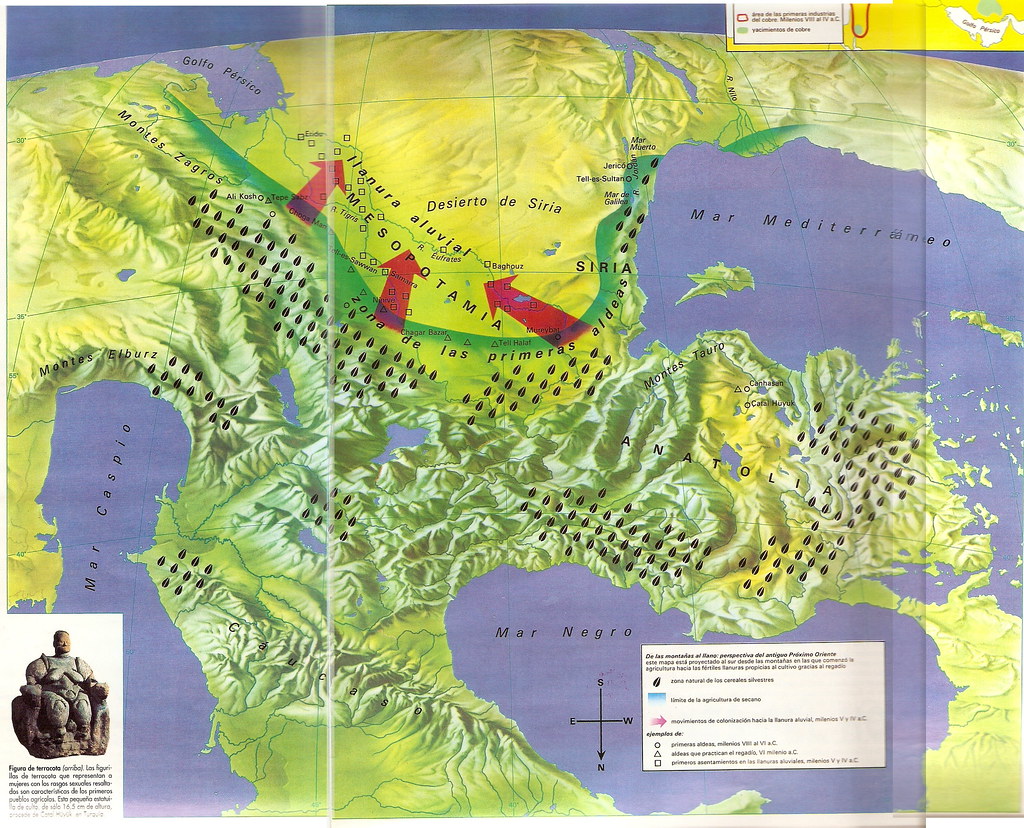
أين ومتى تأصَّلَ قمحُنا الطري؟
رغم أن الأمر، للآن، موضع جدل، لكن، يتفق كثير من الباحثين على حدوثه في منطقة
شرق المتوسط، وقد حصل منذ 11000 عام حيث حصلت موجات ذوبان جليدية بآخر عصر الجليد،
عندها، انتشرت أنواع مختلفة من البقول البرية لتصل الى منطقة الهلال الخصيب. استقرت بهذه المنطقة عدة أنواع من القمح البري:
قمح وحيد الحبة، القمح الأحمر البري، النوع البري Triticum dicoccoides، الدوسر، الشعير العفوي Hordeum spontaneum والشوفان البري Secale vavilovii

منطقة الهلال الخصيب
أحد أهمّ معالم التاريخ البشريّ، هو التحوّل
من مجتمعات التقاط الثمار والصيد المتنقلة الى جماعات مستقرة متخصصة بالزراعة. ففي
غرب آسيا، يُسجّل تحول كهذا عبر ثلاث أنواع من النجيليات البرية، والتي يُعتبر
الانسان من وقتها المتسبب بنهبها المستمر: نوع من القمح البري، القمح
والشعير.
تسجل دراسات جينية كثيرة بأنّ أصل بذرة تدجين تلك
النباتات، أتت من منطقة دياربكر الواقعة جنوب شرق
تركيا الحالية. من هنا، انطلقت بداية زراع " القمح البري الأصغر" عبارة عن
نوع قمح (وحيد الحبّة) ذو جينوم نموذج
AA ومن اقارب نوع القمح الأحمر السلف لقمحنا
الحالي. كما أن تلك المنطقة عرفت البذرة المزروعة من القمح نوع قمح ثنائي الحبّة Triticum dicoccum المعروف بيومنا هذا تحت اسم "النشويّ البريّ".

سنابل نوع القمح البري الأصغر وحيد الحبّة. في الأعلى، سنبلة لنوع قمح مزروع، وفي
الأسفل، سنبلة قمح برّي
وتُعتبر
فلسطين من أوائل الأماكن التي استعمل فيها القمح، حيث يمكننا الحديث عن باقي
"النشويات البرية"، مثل القمح ثنائي الحبّة كسلف مُباشر (أو مباشر تقريبياً) لقمحنا الراهن. رغم أن الإستعمال لا يعني التدجين، فإنّ أوائل الطبقات التي ظهرت فيها تلك الانواع
تعود الى 19000 قبل الميلاد. فكما قلنا، بمجرد حصول تكييفها للزراعة، أعطت المجال
لولادة الزراعة. يظهر تدجين هذا النوع في مناطق الهلال الخصيب اعتباراً من القرن
الحادي عشر قبل الميلاد، بينما يعود ظهورها البرّي الى 13000 عام قبل الميلاد في
الجنوب الشرقي لتركيا الحالية.

مقارنة بين النوع الصُلب الى يسار الصورة، ونوع قمحنا الحديث إلى اليمين
لقد رأينا، للآن، بأنّ الظاهرة الأبرز
لفهم تطور قمحنا، هي تعددية المجموعات الصبغيّة، مع ذلك،
لعب التدجين ورقة هامة أيضاً.
فبخلاف قمحنا الحديث، النشويّ
البريّ عبارة عن نبات ذو بذور صغيرة الحجم، وإنتاجها بكميات قليلة، إضافة
لكونها محمية بواسطة قشرة. ليس هذا فحسب، ككثير من النجيليات فإنّ نوع ثنائي الحبّة عبارة عن نبات، وسط نشره الطبيعي هو الرياح والحيوانات. لهذا، يحتوي
على سنابل متفتحة "هشّة أو مُتقصِّفة"، بحيث تنكسر تلك السنابل بسهولة، بمجرد نضوج
البذور مُحررة سنيبلاتها بصيغة فرديّة. تشكّل تلك السنيبلات وحدة نشر هذا
النبات، كوحدة تشكيل أزهار أساسية في النجيليات، ومن ثمّ تستقبل البذرة. هذا لا
يحصل في قمحنا، في الواقع، سيشكِّل مشكلة.
هل بإمكانكم تخيّل سنبلة
"دون حبوب" وقت الحصاد؟
بالتالي، الأهمّ بالنسبة لنا هو
سنبلة تحتفظ ببذورها بكل لحظة.
مع ذلك، في الطبقات الأقدم،
الموجودة في الجنوب الشرقي من تركيا الحالية وشمال سورية الحالية، والتي تعود الى
10200 عام، تنتمي كل البقايا الأثرية عملياً لسنيبلات متفتحة "هشّة".
لكن، قد تغيَّر الوضع. ففي الطبقات التي تعود الى 7500 عام، فنحن أمام وضع فيه 30%
من السنيبلات من النوع المُنغلق او المُطبق "الغير هشّة". وبمرور 1000
عام، لدى نسبة أكثر من 60% من تلك السنيبلات خصائص كتلك. السنبلة المنغلقة أو
المطبقة، تلك التي تحتفظ بكامل بذورها عند النضوج، فقد أضحت السمة الثابتة
في القمح عملياً.

السنبلة الحديثة المنفتحة (أو "الهشّة") للنشويّ البريّ A ، وسنيبلة منهاB . تفاصيل لمقطع من ذات السنيبلةC .، قمح مدجّن منغلق. D، وتفصيل من عينة عشوائية لسنيبلة منه E ، الرسم البيانيّ F يوضح تناوب عدد من الأحفوريات ذات الصلة لأنواع هشّة منفتحة ولأنواع منغلقة قاسية.طبقات موقع الأسود وطبقات موقع الرماد تنتمي لحالات من الشعير
في الوقت الذي عادت فيه السنابل
منغلقة، مزايا أخرى هي قيد
الانتقاء. سنابل منغلقة مع بذور أكثر صلابة، هو ما يمكننا رؤيته في النشويّ
المزروع، الذي يسميه بعض الباحثين ثنائي الحبّة.
ملمح آخر يميزه عن النشويات، هو حضور
بذور مغطاة ومحمية بقشرة. وتلك ميزة مضبوطة جينياً بما يسمى loci. لكن، تلك
الميزة السلفيّة (من السَلَفْ) تعرضت للإختفاء أيضاً. وبسرعة، تظهر بذور
حرّة من القشرة في طبقات تعود الى 8500 عام.
شكّلت تلك الصيغ النيوليتية السلف
المُباشر لقمحنا الحديث كما "للقمح الصلب" الذي ما يزال يُزرع في شرق
المتوسط وشمال أفريقيا، وهو عبارة عن نبات قد ورث كل الصفات الجيدة (من وجهة نظر زراعية)
التي ظهرت في النشويّ، الى ما أُضِيفَ عبر حضور حبّة عارية، حرّة من القشرة،
الأمر الذي يسهل إستعمالها ومعالجتها.

التعاقب التطوري لنوع قمحنا الحديث،
فمن اليسار الى اليمين نجد: النشويّ البريّ A ، النشويّ المزروع B ، القمح القاسي C والقمح الحديث D ، الأحرف الموجودة في الأسفل، تمثل جينات تلك
الأنواع
بالنهاية، بلغ تدجين القمح ذروته
بظهور نوع القمح الطري. الذي كان نتاج تعدد تنوعات نوع القمح البرّي Triticum turgidum من بذور عارية وسنابل منغلقة في الغالب، ومن خلال " عشبة الماعز " تلك المسماة دوسر شبه مكنسي. الأمر الذي يُعطي الأصل لقمحنا الحديث، كنوع خاص حديث جداً. رغم عدم تناسق عمره بكل حساباته، فإنّ الحساب على قاعدة ساعاته
الجزيئية، الذي أجراه بعض الباحثين، ساعدهم على تحديد عمره بحدود 9500 قبل الميلاد، بينما يُرجعه
باحثون آخرون لزمن أبكر لحوالي 6000 عام قبل الميلاد، فيربطونه ببدء الزراعة جنوب
غرب بحر قزوين، المكان الذي تنتشر فيه "عشبة الماعز" تلك بصورة
طبيعية.
تمّ تبرير نجاح نوع القمح الطري تقليدياً بمواجهة "القمح القاسي" عبر الإستعانة
بصلابته العالية، الأمر الذي شكّل إفتراضاً أعرجاً! حيث يُنتج "القمح القاسي" بذور أكبر حجماً وذو قدرة على إنتاج مشابه بشروط نمو مفضلة.
من وقت قريب
جداً، طرح باحثون آخرون مقارنة لأنواع قمح طري تعكس مزاياه الهامّة، مثل تمتعه بلدونة عالية بمراحل مختلفة وشروط
الاستبراد (معالجة البذور بالبرد بُغية تسريع انتاشها او انباتها وتسريع نموّ
النبات وازهاره .. قاموس المورد اسباني عربي)، سامحة له بالنمو والإنتاج تحت
تأثير ظروف مُناخية مختلفة وتطوير مقاومة للملوحة وانخفاض قيمة pH لحضور
الالمنيوم وللتبريد ومقاومة عالية للأوبئة والأمراض؛ كذلك امتلاك قدرة عالية لتحضير نماذج مختلفة من المنتجات الغذائية وهو الأهمّ.
تطور القمح. لاحظوا التغيرات في شكل السنبلة
المصدر الإسباني:
ليست هناك تعليقات:
إرسال تعليق